Molekulare Spuren von Lernen und Gedächtnis
Forschungsbericht (importiert) 2019 - Max-Planck-Institut für Hirnforschung
Lernen auf Zellebene
Lernen und Gedächtnis sind täglich benötigte Leistungen unseres Gehirns. Paradoxerweise muss das selber aus instabilen Proteinen aufgebaute Gehirn dazu einerseits veränderbar sein, gleichzeitig aber auch in der Lage, Informationen für lange Zeit stabil zu bewahren. Die Grundeinheiten des Gehirns sind Nervenzellen (Neuronen) – polar gebaute Zellen, die außer dem Zellkörper lange Fortsätze zum Senden (Axone) und Empfangen (Dendriten) von Signalen haben. Die kleinste Ebene, auf der Vorgänge zu Lernen und Gedächtnis nachgewiesen wurden, sind die Kontakt- und Kommunikationspunkte zwischen Axonen und Dendriten verschiedener Neuronen, die sogenannten Synapsen (Abb. 1). Ein Neuron leitet seine Information zunächst als elektrisches Signal entlang des Axons. Am sendenden Teil der Kontaktstelle, der Präsynapse, wird das elektrische Signal in ein chemisches umgewandelt. Hierzu wird ein Botenstoff, der erregend oder hemmend sein kann, aus kleinen Vesikeln – quasi Botenstoff- Containern – freigesetzt. Der Empfänger-Teil der Synapse gehört zu einem anderen Neuron und wird Postsynapse genannt. Hier bindet der Botenstoff an einen Rezeptor und löst erneut ein elektrisches Signal aus.
. Die Kommunikationsstellen zwischen Nervenzellen nennt man Synapsen. Ein Dendrit kann tausende von Eingangsstellen (Postsynapsen) für Informationen aus Senderstellen (Präsynapsen) von Axonen anderer Neurone besitzen. An der Präsynapse wird ein ankommendes elektrisches Signal in ein chemisches umgewandelt: Mit Botenstoff gefüllte Vesikel schütten diesen Botenstoff aus, der an der Postsynapse an Rezeptoren bindet und dort ein elektrisches Signal in der nächsten Zelle auslöst. Lokale Prozesse wie der Auf- und Abbau von Proteinen in der Nähe von Synapsen können erklären, wie einzelne Synapsen unabhängig von den anderen ihre Übertragungsstärke ändern können. Früher nahm man an, dass Proteinsynthese nur im Zellkörper nahe des Zellkerns stattfindet, da mRNAs - die Baupläne für Proteine - im Zellkern hergestellt werden.")
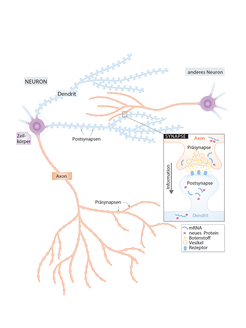
Abb.1: Organisation und Kommunikation einer Nervenzelle. Neuronen haben im Gegensatz zu anderen Zellen zusätzlich zum Zellkörper lange Ausläufer (Axon und Dendriten). Die Kommunikationsstellen zwischen Nervenzellen nennt man Synapsen. Ein Dendrit kann tausende von Eingangsstellen (Postsynapsen) für Informationen aus Senderstellen (Präsynapsen) von Axonen anderer Neurone besitzen. An der Präsynapse wird ein ankommendes elektrisches Signal in ein chemisches umgewandelt: Mit Botenstoff gefüllte Vesikel schütten diesen Botenstoff aus, der an der Postsynapse an Rezeptoren bindet und dort ein elektrisches Signal in der nächsten Zelle auslöst. Lokale Prozesse wie der Auf- und Abbau von Proteinen in der Nähe von Synapsen können erklären, wie einzelne Synapsen unabhängig von den anderen ihre Übertragungsstärke ändern können. Früher nahm man an, dass Proteinsynthese nur im Zellkörper nahe des Zellkerns stattfindet, da mRNAs - die Baupläne für Proteine - im Zellkern hergestellt werden.
Ein Neuron hat tausende von Synapsen, über die es Signale von anderen Neuronen erhält. Die Fähigkeit von Synapsen, unabhängig von anderen Synapsen des gleichen Neurons langfristig ihre Übertragungsstärke zu verändern, gilt als Lernen und Gedächtnis auf Zellebene. Die Produktion neuer Proteine, den Arbeitsmaschinen einer Zelle, spielt dabei eine entscheidende Rolle. Sowohl Lernen als auch das langfristige Verstellen der Übertragungsstärke an Synapsen funktionieren nicht mehr, wenn die Proteinproduktion verhindert wird. Doch wie kann ein Neuron eine Kontaktstelle unter tausenden gezielt verändern? Wie kennen die Proteine ihren Bestimmungsort?
Eine unmittelbar einleuchtende Erklärung lieferte der Nachweis, dass neue Proteine auf Aktivitätssignale hin dezentral, also lokal nahe an Synapsen, produziert werden können [1]. Normalerweise werden Proteine anhand von zentral im Zellkörper erstellten Bauanleitungen, den mRNAs, auch zentral im Zellkörper erzeugt und danach in der Zelle verteilt. Wir konnten in Neuronen erstaunlich viele mRNAs in Synapsen-Nähe nachweisen. Doch was sind die Details lokaler Proteinproduktion? Bildlich gesprochen: Heißt ,lokale‘ Produktion, dass z. B. für Hessen produziert wird oder nur für Frankfurt? Wo genau steht die dezentrale Fabrik? Welche Produkte werden zu einem bestimmten Zeitpunkt gefertigt und gibt es eine Aufgabenteilung oder Spezialisierung in unterschiedlichen Regionen? Woher kommt die Energie?
Spezialisierung in der lokalen Proteinproduktion
Die Frage ‚wo genau‘ beantwortet man in der Wissenschaft in der Regel über Mikroskopie - in unserem Falle eine Herausforderung, da die Distanzen zwischen denkbaren Produktions-Orten an der Auflösungsgrenze der Lichtmikroskopie liegen. Zudem benötigten wir Methoden, die Proteine als ‚neu‘ identifizieren. Letztere entwickeln wir seit Jahren [1;2;3] und kombinierten sie mit einem Trick: Neuronen wurden in eine Matrix eingebettet, die ähnlich einer Windel, Feuchtigkeit aufnehmen und quellen kann. Verankert man vor dem Quellvorgang die Proteine chemisch an der Matrix, kann man unter geeigneten Quellbedingungen alle ursprünglichen Strukturen proportional vergrößern. Man erhält damit ca. 4fach vergrößertes Gewebe (Abb. 2 A), womit die Auflösungsgrenze der Lichtmikroskopie überwunden ist.
 Ein Gewebeschnitt vor und nach dem Quellen zeigt, dass er tatsächlich real größer wird. (B) Unterschiedliche Substanzen (X,Y,Z) lösen 3 verschiedene Arten zellulären Lernens aus, die rechts oben genannt sind (Potenzierung = Verstärkung, Depression = Abschwächung der Informationsübertragung). Nach Behandlung mit diesen Substanzen, Markierung neu produzierter Proteine und Expansion konnten wir mikroskopisch untersuchen, wo genau die Proteinproduktion unter den jeweiligen Bedingungen hoch reguliert wurde (grüne Kästchen) und wo nicht (weiße Kästchen). Es ergab sich ein eindeutiger Code. (C) Ausschnitte aus einem Film mit gentechnisch markierten Mitochondrien zeigen, dass ein großer Mitochondrien-Cluster in einem Dendriten (grüner Pfeil) stabil an einem Ort bleibt. Im Axon sind die Mitochondrien kleiner und bewegen sich oft schnell entlang des Ausläufers (rote Pfeile). (D) Aktiviert man einzelne Postsynapsen (*) in einem bestimmten Muster, dann ‚lernen‘ diese, dass sie stärker antworten müssen. Sie nehmen dabei an Größe zu. Legt man einzelne Mitochondrien-Cluster als lokale Energieversorger lahm (hier im Bereich der gestrichelten Box), dann findet dieses Größenwachstum und Lernen im Bereich der abgeschalteten Energieversorgung nicht statt.")
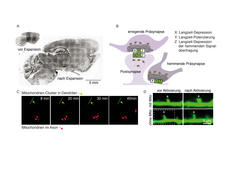
Abb.2: Effekt und Ergebnis der Expansionsstrategie/ Mitochondrien als lokale Energieversorger in Neuronen.
(A) Ein Gewebeschnitt vor und nach dem Quellen zeigt, dass er tatsächlich real größer wird. (B) Unterschiedliche Substanzen (X,Y,Z) lösen 3 verschiedene Arten zellulären Lernens aus, die rechts oben genannt sind (Potenzierung = Verstärkung, Depression = Abschwächung der Informationsübertragung). Nach Behandlung mit diesen Substanzen, Markierung neu produzierter Proteine und Expansion konnten wir mikroskopisch untersuchen, wo genau die Proteinproduktion unter den jeweiligen Bedingungen hoch reguliert wurde (grüne Kästchen) und wo nicht (weiße Kästchen). Es ergab sich ein eindeutiger Code. (C) Ausschnitte aus einem Film mit gentechnisch markierten Mitochondrien zeigen, dass ein großer Mitochondrien-Cluster in einem Dendriten (grüner Pfeil) stabil an einem Ort bleibt. Im Axon sind die Mitochondrien kleiner und bewegen sich oft schnell entlang des Ausläufers (rote Pfeile). (D) Aktiviert man einzelne Postsynapsen (*) in einem bestimmten Muster, dann ‚lernen‘ diese, dass sie stärker antworten müssen. Sie nehmen dabei an Größe zu. Legt man einzelne Mitochondrien-Cluster als lokale Energieversorger lahm (hier im Bereich der gestrichelten Box), dann findet dieses Größenwachstum und Lernen im Bereich der abgeschalteten Energieversorgung nicht statt.
Behandelten wir Neuronen vorher mit Substanzen, die verschiedene Arten zellulären Lernens auslösten, ließ sich nun sagen, ob die Proteinproduktion im Sender- oder Empfänger-Teil der Synapse stattfand. Hemmende und erregende Synapsen unterschieden sich. Erstaunlicherweise fanden wir eine Art Code aus Proteinproduktions-Orten, der verschiedene Arten des Zell-Lernens wiederspiegelte (Abb. 2; [4]).
Dass in ausgereiften Synapsen - nur diese zeigen Zell-Lernen - präsynaptisch Proteine produziert werden, wurde bisher angezweifelt. Um diesem aufregenden Befund nachzugehen, reinigten wir biochemisch die winzigen Sender-Kompartimente von erregenden Neuronen auf. Aus diesen isolierten wir die mRNA und zeigten, dass hier tatsächlich ein spezieller Satz von Bauanleitungen zu finden ist: vor allem für Proteine, die den Ausschüttungsort der chemischen Botenstoffe organisieren.
Eine Frage der Energieversorgung
Wie die moderne Gesellschaft benötigt auch ein Neuron große Mengen an Energie, für die unterschiedliche Wege der Gewinnung existieren. Die Frage nach der Energieversorgung für zelluläre Lernvorgänge führte uns zu den Mitochondrien, den Kraftwerken der Zelle [5]. Gentechnisch markiert und live am Mikroskop verfolgt, verhielten sich Mitochondrien in Axonen und Dendriten völlig verschieden. In Dendriten sind Mitochondrien als Cluster von ~30 µm Ausdehnung lange stabil an einem Ort zu finden, in Axonen bilden sie dagegen kleine, mobile Pakete (Abb. 2 C).
Wir simulierten einen Ausfall der lokalen Energieversorgung, indem wir den Farbstoff „Killerred“ an Mitochondrien lenkten. Wird er mittels Laserlicht aktiviert, zerstört er seine unmittelbare Umgebung. Wir konnten so gezielt einzelne Kraftwerk-Cluster außer Gefecht setzen. Der lokale Ausfall beeinflusste unter Normalbedingungen die Proteinproduktion nicht. Versuchten wir jedoch zelluläres Lernen an Synapsen im Bereich der lahmgelegten Kraftwerke zu induzieren, konnte die dafür notwendige Proteinproduktion genau in diesem Bereich nicht mehr hochgefahren werden. Die Folge war, dass mit dem Lernvorgang einhergehende Strukturveränderungen nicht mehr nachhaltig manifestiert wurden und kein Lernen erfolgte (Abb. 2 D). Die Klärung dieser Details ist auch deshalb von Interesse, weil Fehlfunktionen sowohl von Mitochondrien als auch bei der Proteinproduktion ein Charakteristikum neurodegenerativer Erkrankungen sind. Wichtig werden diese Erkenntnisse dann, wenn Prozesse aus der Bahn laufen und die Herausforderung ist, Ursache von Wirkung zu unterscheiden und diese Fehlfunktionen zu korrigieren.
Literaturhinweise
DOI: 10.15252/embr.201744045
DOI:10.1038/nmeth.3319
Nature Biotechnology 35, 1196-1201 (2017)
DOI: 10.1038/nbt.4016
DOI: 10.1016/j.cell.2018.12013
DOI: 10.1126/science.aau3644