Forschungsbericht (importiert) 2007 - Max-Planck-Institut für Hirnforschung
Autoren
Schmidt, Kerstin E.
Abteilungen
Kortikale Funktion und Dynamik (Dr. Kerstin Schmidt) MPI für Hirnforschung, Frankfurt/Main
Zusammenfassung
Nervenzellen in der primären Sehrinde wurden für Analyseeinheiten winziger Gesichtsfeldausschnitte (rezeptive Felder) gehalten. Heute ist bekannt, dass die neuronale Antwort durch den Kontext, in dem der optimale Reiz präsentiert wird, moduliert wird. Mittels optischer Ableitung wird untersucht, wie komplexe Sehreize in Populationsantworten integriert werden und welche neuronalen Strukturen die unterschwelligen modulierenden Einflüsse vermitteln.
Rezeptive Felder und visuelle Stimuli
Einzelne Nervenzellen in sensorischen Hirnrindenarealen reagieren auf Eindrücke aus örtlich begrenzten Ausschnitten der Sinneswelt (rezeptive Felder) mit einer Erhöhung ihrer Aktionspotenzialrate. In der primären Sehrinde werden auf diese Art visuelle Reize in kleinen Feldern mit weniger als 1° Sehwinkel bearbeitet. In höheren Sehrindenarealen vergrößern sich die rezeptiven Felder zunehmend, die bevorzugten Reize werden komplexer, und einzelne Zellen antworten beispielsweise selektiv auf Gesichter. Obwohl in der anatomischen Verbindungsstruktur zwischen den Arealen, die mit dem Sehvorgang befasst sind, ein hierarchischer Aufbau existiert, befindet sich nirgendwo im Hirn ein Sehareal, in welchem die gesamte visuelle Information zusammenläuft. Vielmehr bestehen parallele Verarbeitungspfade, eine ausgeprägte Quervernetzung und rückwärts gerichtete Projektionen (Feedback) nebeneinander. Seit einigen Jahren ist jedoch bekannt, dass Neurone in der primären Sehrinde nicht ausschließlich Zugang zu Reizen innerhalb ihres rezeptiven Feldes haben. Sie besitzen zusätzlich ein modulierendes Umfeld, welches wesentlich größer als ihr klassisches rezeptives Feld ist. Visuelle Stimuli innerhalb dieses Umfeldes allein lösen zwar kein Aktionspotenzial an der betreffenden Nervenzelle aus, können aber die Antwortstärke auf Stimuli, die im klassischen rezeptiven Feld präsentiert werden, modulieren. Da nebeneinander liegende Neurone benachbarte Punkte des Gesichtsfeldes bearbeiten, fließt über unterschwellige Schwankungen des Membranpotenzials der visuelle Kontext so mit ein.
Dieser Bericht handelt von Untersuchungen darüber, in welcher Art Sehreize außerhalb des klassischen rezeptiven Feldes auf der Ebene von Neuronenpopulationen integriert werden und welche neuronalen Schaltkreise für die Weitergabe unterschwelliger Einflüsse verantwortlich sind. Zur Darstellung von Neuronenpopulationen eignet sich die optische Ableitung intrinsischer Signale, das heißt lokaler Schwankungen des Blutsauerstoffgehalts und des Blutdurchflusses, die mit Veränderungen sowohl unter- als auch überschwelliger neuronaler Aktivität korreliert sind [1].
Lokale und globale Richtungsselektivität in der primären Sehrinde – Plaid Stimuli
Nervenzellen in der primären Sehrinde antworten bevorzugt auf lineare Bewegungen von Kontrastgrenzen. Dabei reagieren sie selektiv auf die Orientierung des jeweiligen Liniensegments und meist auch auf eine der beiden dazu orthogonal verlaufenden Bewegungsrichtungen. Aufgrund ihres kleinen „klassischen“ rezeptiven Feldes ist für die Nervenzellen aber die globale physikalische Bewegungsrichtung einer geraden Linie unterspezifiziert. Eine ganze Familie von Bewegungsrichtungen, welche nicht orthogonal zur Kontrastgrenze verlaufen, ist für einen lokalen Bewegungsdetektor nicht von den orthogonal verlaufenden zu unterscheiden. Schaut man beispielsweise durch ein Schlüsselloch, vor dem sich eine lange Linie parallel zu ihrer Orientierung bewegt, nimmt der Beobachter in diesem Extremfall sogar gar keine Bewegung mehr war, solange nicht ein Ende des Liniensegmentes die Öffnung passiert.
Lange Zeit ging man daher davon aus, dass Zellen in primären Sehrindenarealen aufgrund dieser Einschränkung die echte Bewegungsrichtung eines komplexen Musters oder Objektes nicht anzeigen können, und dies wurde auch mehrfach durch Einzelzellableitungen bei Primaten und Katzen bestätigt. Es wurde ein Zweistufenmodell für Bewegungserkennung aufgestellt, nach dem in frühen Sehrindenarealen zunächst die Bewegungsrichtungen lokaler Komponenten ausgewertet werden, um dann in höheren Arealen mit größeren rezeptiven Feldern die globale Bewegungsrichtung des Gesamtobjekts zu rekonstruieren. Der visuelle Stimulus, der entwickelt wurde, um lokale und globale Bewegungsrichtung gegeneinander abzuwägen, besteht aus zwei übereinander gelegten Balkengittern, die sich in zwei verschiedene Richtungen bewegen (Abb. 1). Durch Variation des Kontrastes an den jeweiligen Schnittstellen der beiden Gitter nimmt der Betrachter entweder tatsächlich zwei sich übereinander schiebende Gitter wahr oder aber nur ein einziges Karomuster (Plaid), welches sich in diejenige Richtung bewegt, die genau zwischen den Bewegungsrichtungen der beiden Gitterkomponenten liegt. Die Varianten können besonders bei niedrigem Kontrast auch miteinander abwechseln, obwohl physikalisch derselbe Reiz dargeboten wird [2].
Im primären visuellen Kortex anästhesierter Katzen wurde untersucht, ob sich die intrinsischen Aktivitätsverteilungen durch Plaid-Stimulation bei verschiedenen Schnittstellenkontrasten unterscheiden. Es stellte sich heraus, dass Plaids, welche in der Regel die Wahrnehmung eines sich in eine einzige Richtung bewegenden Karomusters hervorrufen, Aktivitätsverteilungen erzeugen, die denjenigen ähneln, die durch ein einziges in die Musterrichtung bewegtes Balkengitter erzeugt werden, obwohl sich die Plaid-Komponenten in ganz andere Richtungen bewegen. Plaid Stimuli hingegen, die so konstruiert sind, dass bevorzugt zwei sich übereinander schiebende Ebenen wahrgenommen werden, erzeugen Aktivitätsverteilungen, die denen der einzelnen Komponentengitter gleich kommen (Abb. 1). Daraus lässt sich schließen, dass schon in der primären Sehrinde zwischen globaler Musterrichtung und lokalen Bewegungskomponenten auf Populationsebene unterschieden wird. Dies ist sehr bemerkenswert, da rezeptive Felder auf dieser Verarbeitungsebene in der Regel nur ein Liniensegment abdecken. Da bei Einzelzellableitungen hier Muster-richtungsselektive Neurone meist nicht oder nur vereinzelt in wachen Tieren beobachtet wurden, sprechen unsere Ergebnisse dafür, dass die Musterinformation in der unterschwelligen Aktivität von außerhalb des klassischen rezeptiven Feldes enthalten ist [3].
Optisch abgeleitete Aktivitätsverteilungen im primären visuellen Kortex bei Stimulation durch Plaid-Stimuli. Plaids sind aus zwei in Richtung der Pfeile bewegten Balkengittern zusammengesetzt und werden entweder als zwei sich übereinander schiebende Ebenen oder als ein nach oben driftendes Gesamtmuster wahrgenommen. Als Reize dienten zwei Plaids (A, E, B, F), ihre beiden Komponenten (G, H) und ein Modellstimulus für ihre Musterbewegung (C). Aktivitätskarten wurden entweder durch die Summe der beiden Komponentenaktivierungen (erste Spalte) oder die Mustermodelaktivität (zweite Spalte) dividiert. Ein Plaid mit niedrigem Schnittstellenkontrast (A, E), bevorzugt als zwei Ebenen wahrgenommen, ruft Aktivitätsverteilungen hervor, die durch die Summe der beiden Komponentenaktivierungen erklärt werden können (flache Modulation in A, starke komponentenähnliche Modulation in E, vergleiche E, G, H). Ein Plaid mit hohem Schnittstellenkontrast wird meist als sich in die Gesamtrichtung bewegendes Muster wahrgenommen. Tatsächlich aktiviert das Plaid in der primären Sehrinde Domänen, die denen des Mustermodells ähneln (starke Musteraktivierung in B, flache komponentenähnliche Modulation in F, vergleiche B, C), obgleich einzelne Neurone nur kleine Ausschnitte des Gesamtplaids bearbeiten. D: Oberflächenbild des abgeleiteten Gebiets. Rote Sterne in den Mustermodelldomänen (C) dienen der Orientierung. Die Antwortstärke ist als Grauwert kodiert.
Optisch abgeleitete Aktivitätsverteilungen im primären visuellen Kortex bei Stimulation durch Plaid-Stimuli. Plaids sind aus zwei in Richtung der Pfeile bewegten Balkengittern zusammengesetzt und werden entweder als zwei sich übereinander schiebende Ebenen oder als ein nach oben driftendes Gesamtmuster wahrgenommen. Als Reize dienten zwei Plaids (A, E, B, F), ihre beiden Komponenten (G, H) und ein Modellstimulus für ihre Musterbewegung (C). Aktivitätskarten wurden entweder durch die Summe der beiden Komponentenaktivierungen (erste Spalte) oder die Mustermodelaktivität (zweite Spalte) dividiert. Ein Plaid mit niedrigem Schnittstellenkontrast (A, E), bevorzugt als zwei Ebenen wahrgenommen, ruft Aktivitätsverteilungen hervor, die durch die Summe der beiden Komponentenaktivierungen erklärt werden können (flache Modulation in A, starke komponentenähnliche Modulation in E, vergleiche E, G, H). Ein Plaid mit hohem Schnittstellenkontrast wird meist als sich in die Gesamtrichtung bewegendes Muster wahrgenommen. Tatsächlich aktiviert das Plaid in der primären Sehrinde Domänen, die denen des Mustermodells ähneln (starke Musteraktivierung in B, flache komponentenähnliche Modulation in F, vergleiche B, C), obgleich einzelne Neurone nur kleine Ausschnitte des Gesamtplaids bearbeiten. D: Oberflächenbild des abgeleiteten Gebiets. Rote Sterne in den Mustermodelldomänen (C) dienen der Orientierung. Die Antwortstärke ist als Grauwert kodiert.
Der Einfluss von Feedback auf globale Richtungsselektivität
Muster-richtungsselektive Neurone fand man häufiger in einem höheren visuellen Areal des parietalen Pfades, der auf die Verarbeitung von Bewegungsinformation spezialisiert ist. Bei der Katze liegt dieses Areal im postero-medialen suprasylvischen Sulkus (pMS). Tatsächlich weist pMS große richtungsselektive Felder auf und projiziert stark zurück in die primären Sehrindeareale (Area 17/18). Um die Herkunft der Muster-richtungsselektiven Aktivität in der primären Sehrinde zu untersuchen, wurde dieses Areal während der Plaid-Stimulation durch Kühlung reversibel ausgeschaltet. Es zeigte sich, dass bei Deaktivierung der Feedbackprojektionen aus pMS die Unterscheidung zwischen Musterrichtung und lokaler Komponentenrichtung in der primären Sehrinde (Area17) verloren geht. Die Muster-richtungsselektiven Aktivierungen, die durch das kontrastreiche Plaid hervorgerufen wurden, gleichen nun denjenigen, die durch die kontrastarme 2-Ebenen-Variante hervorgerufen werden und nicht mehr der durch das in die Musterrichtung bewegte Balkengitter hervorgerufenen Populationsantwort (Abb. 2). Dies ist der erste experimentelle Hinweis darauf, dass globale Richtungsselektivität durch Feedback an die primäre Sehrinde vermittelt werden könnte [4].
Plaid-Aktivität bei Kühlungsdeaktivierung des Areals pMS, welches große bewegungssensitive rezeptive Felder besitzt und Rückprojektionen in die primäre Sehrinde sendet. A: Farbkodierte Orientierungskarte und Oberflächenbild des abgeleiteten Gebiets. Die bevorzugte Stimulusorientierung ist für jeden Pixel gemäß dem Farbschema farbkodiert. B: Auf die Farbkarte überlagerte Aktivitätsverteilungen aus Abb. 1 in der Kontrollphase (schwarz), bei pMS-Kühlung (blau) und nach Abschalten der Kühlung (grau). Je stärker die Aktivierung, desto heller die Farbe. Es fällt auf, dass im Gegensatz zum schwach kontrastierten Plaid (erste Spalte) das Plaid mit hohem Schnittstellenkontrast (zweite Spalte) die gleichen Domänen (gelb) aktiviert wie das Mustermodell (dritte Spalte). Bei pMS-Kühlung nimmt die Musteraktivität für dieses Plaid ab, wohingegen sie für das Modell erhalten bleibt. Dafür nimmt die komponentenähnliche Aktivität zu. Alle Effekte verschwinden bei Erwärmung von pMS wieder.
Plaid-Aktivität bei Kühlungsdeaktivierung des Areals pMS, welches große bewegungssensitive rezeptive Felder besitzt und Rückprojektionen in die primäre Sehrinde sendet. A: Farbkodierte Orientierungskarte und Oberflächenbild des abgeleiteten Gebiets. Die bevorzugte Stimulusorientierung ist für jeden Pixel gemäß dem Farbschema farbkodiert. B: Auf die Farbkarte überlagerte Aktivitätsverteilungen aus Abb. 1 in der Kontrollphase (schwarz), bei pMS-Kühlung (blau) und nach Abschalten der Kühlung (grau). Je stärker die Aktivierung, desto heller die Farbe. Es fällt auf, dass im Gegensatz zum schwach kontrastierten Plaid (erste Spalte) das Plaid mit hohem Schnittstellenkontrast (zweite Spalte) die gleichen Domänen (gelb) aktiviert wie das Mustermodell (dritte Spalte). Bei pMS-Kühlung nimmt die Musteraktivität für dieses Plaid ab, wohingegen sie für das Modell erhalten bleibt. Dafür nimmt die komponentenähnliche Aktivität zu. Alle Effekte verschwinden bei Erwärmung von pMS wieder.
Modulation unterschwelliger Orientierungsselektivität durch Feedback
Die durch einen örtlich begrenzten Sehreiz hervorgerufene Modulation des intrinsischen Signals nimmt einen Kortexbereich ein, der um ein Vielfaches größer ist als der Bereich, der mit einer Erhöhung der Aktionspotenzialrate, also überschwellig, reagiert. Dabei besitzt die unterschwellig durch den Kontext modulierte Aktivität die gleiche Orientierungsselektivität wie die Aktionspotenziale im Zentrum der überschwellig gereizten Region. Das führt dazu, dass räumlich komplementäre Reize bei optischer Ableitung überlappende Erregungsmuster produzieren (Abb. 3). Ein Teil der orientierungsselektiven Domänen, die durch Stimulation mit einem bewegten Balkengitter mit einer zentralen Lücke (Umfeldstimulus) angesprochen werden, antwortet auch auf ein kleines Balkengitter in der Größe der Lücke (Feldstimulus), obwohl die beiden Sehreize selbst nicht überlappen.
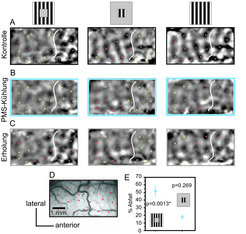
Optische Aktivität bei Stimulation mit komplementären Balkengittern (Umfeld, Feld) und Ganzfeld (dritte Spalte) vertikaler Orientierung während Kühlung des Areals pMS. Sterne kennzeichnen die Zentren der aktiven Domänen bei Ganzfeldstimulation in der Kontrollbedingung (oben rechts, rot: Überlappung mit der Ganzfeldkontrolle, gelb: Nichtüberlappung). A: Aktive Domänen in der Kontrollbedingung. Erste Spalte: Umfeldaktivierung vornehmlich anterior, zweite Spalte: Feldaktivierung vornehmlich posterior. Trotz nicht überlappender Sehreize werden einige Domänen von sowohl Umfeld- als auch Feldstimuli angesprochen (rote Sterne links der weißen Linie). B: Bei pMS-Deaktivierung nimmt die Antwortstärke für alle Reize ab. Dabei schrumpfen die vom Umfeld modulierten Bereiche sowie die Feld/Umfeld-Überlappungszone. C: Erholungsphase. D: Kortexoberfläche. E: Kühleffekte wirken sich stärker aus auf Umfeld- als auf Feldreizung: Der prozentuale Abfall der Übereinstimmung mit der Ganzfeldkontrolle ist bei Umfeldreizung signifikant (n=13). Die Antwortstärke ist als Grauwert kodiert.
Optische Aktivität bei Stimulation mit komplementären Balkengittern (Umfeld, Feld) und Ganzfeld (dritte Spalte) vertikaler Orientierung während Kühlung des Areals pMS. Sterne kennzeichnen die Zentren der aktiven Domänen bei Ganzfeldstimulation in der Kontrollbedingung (oben rechts, rot: Überlappung mit der Ganzfeldkontrolle, gelb: Nichtüberlappung). A: Aktive Domänen in der Kontrollbedingung. Erste Spalte: Umfeldaktivierung vornehmlich anterior, zweite Spalte: Feldaktivierung vornehmlich posterior. Trotz nicht überlappender Sehreize werden einige Domänen von sowohl Umfeld- als auch Feldstimuli angesprochen (rote Sterne links der weißen Linie). B: Bei pMS-Deaktivierung nimmt die Antwortstärke für alle Reize ab. Dabei schrumpfen die vom Umfeld modulierten Bereiche sowie die Feld/Umfeld-Überlappungszone. C: Erholungsphase. D: Kortexoberfläche. E: Kühleffekte wirken sich stärker aus auf Umfeld- als auf Feldreizung: Der prozentuale Abfall der Übereinstimmung mit der Ganzfeldkontrolle ist bei Umfeldreizung signifikant (n=13). Die Antwortstärke ist als Grauwert kodiert.
Lange Zeit wurden nahezu ausschließlich die langen Horizontalverbindungen innerhalb der primären Sehrinde mit diesen Beobachtungen in Verbindung gebracht, weil sie Nervenzellen ähnlicher Orientierungsselektivität miteinander verknüpfen. Aber auch die Topographie der Feedbackverbindungen ermöglicht eine spezifische laterale Ausbreitung unterschwelliger Erregung. So stellte sich bei Ausschaltung der Feedbackverbindungen von pMS heraus, dass die von Umfeld- und Feldstimulus modulierten Kortexbereiche schrumpfen, besonders in den überlappend antwortenden Regionen und insbesondere für den mehr Kontext einschließenden Umfeldstimulus. Dies deutet darauf hin, dass Feedbackprojektionen einen stabilisierenden Effekt auf räumliche Stimulusrepräsentationen haben [5]. Zusammen mit den Ergebnissen zur globalen Richtungsselektivität liegt die Vermutung nahe, dass globale Stimuluseigenschaften in höheren Seharealen integriert und als unterschwellige Aktivität von außerhalb des klassischen rezeptiven Feldes an primäre Sehareale zurückgesendet werden.
Modulation unterschwelliger Orientierungsselektivität durch callosale Verbindungen
Es gibt keinen der reversiblen Kortexkühlung vergleichbaren Ansatz, die Horizontalverbindungen innerhalb eines Areals auszuschalten. Es ist jedoch anzunehmen, dass Verbindungen zwischen homologen Arealen in den beiden Hirnhälften ähnliche Aufgaben erfüllen müssen wie die Horizontalverbindungen. Das Gesichtsfeld wird an der vertikalen Mittellinie geteilt und je eine Hälfte in die gegenüberliegende Sehrinde gesandt. Die beiden Sehrinden sind durch einen dicken Faserstrang, das Corpus Callosum, miteinander verbunden. Nervenzellen mit rezeptiven Feldern direkt an der Mittellinie beziehen oft den ipsilateralen Anteil ihres Feldes über callosale Verbindungen. Die Fasern sind aber keineswegs auf die kortikale Repräsentation der Mittellinie, welche typischerweise an den Arealgrenzen lokalisiert ist, beschränkt, sondern finden sich dort nur in ihrer größten Dichte. In der Nachwuchsgruppe wurde der Einfluss callosaler Verbindungen auf die von Feld- und Umfeldstimulus hervorgerufenen kortikalen Aktivierungen in den beiden frühen Sehrindenarealen 17 und 18 getestet. Es stellte sich heraus, dass bei Kühlung des gleichen Gebiets in der gegenüberliegenden Hirnhälfte zusätzlich zu einem unspezifischen Abfall der Antwortselektivität die Stimulus-modulierten Kortexbereiche schrumpfen (Abb. 4), und zwar umso stärker, je näher der visuelle Reiz an die vertikale Mittellinie heranreicht. Dabei ist auch hier in der Regel der größere Umfeld-Stimulus stärker betroffen [6].
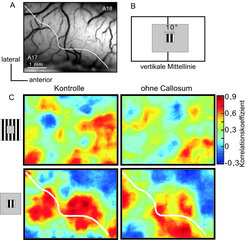
Optische Aktivität bei Stimulation mit komplementären Balkengittern während Kühlung des gleichen Areals in der gegenüberliegenden Hirnhälfte. Die Kühlung schaltet Eingänge über das Corpus Callosum, die für die Kontinuität des Bildes über die vertikale Mittellinie verantwortlich gemacht werden, aus. A: Oberfläche des abgeleiteten Areals; weiße Linie: Area 17/18-Grenze und kortikale Repräsentation der Gesichtsfeldmitte. B: Der Feldstimulus wurde direkt an die Mittellinie positioniert. C: Korrelationskarten für Feld- und Umfeldstimulus. An jedem Punkt der Karte wurde die tatsächliche Aktivitätsverteilung mit einer für den jeweiligen Stimulus zu erwartenden Idealverteilung korreliert. Der Feldstimulus ist sowohl in Area 17 als auch 18 abgebildet. In der Kontrollbedingung konfluieren die beiden Aktivitätsschwerpunkte an der Mittellinie. Bei Ausschaltung callosaler Eingänge nehmen die Korrelationen mit der Idealverteilung ab und die Repräsentationen in den beiden Arealen schrumpfen.
Optische Aktivität bei Stimulation mit komplementären Balkengittern während Kühlung des gleichen Areals in der gegenüberliegenden Hirnhälfte. Die Kühlung schaltet Eingänge über das Corpus Callosum, die für die Kontinuität des Bildes über die vertikale Mittellinie verantwortlich gemacht werden, aus. A: Oberfläche des abgeleiteten Areals; weiße Linie: Area 17/18-Grenze und kortikale Repräsentation der Gesichtsfeldmitte. B: Der Feldstimulus wurde direkt an die Mittellinie positioniert. C: Korrelationskarten für Feld- und Umfeldstimulus. An jedem Punkt der Karte wurde die tatsächliche Aktivitätsverteilung mit einer für den jeweiligen Stimulus zu erwartenden Idealverteilung korreliert. Der Feldstimulus ist sowohl in Area 17 als auch 18 abgebildet. In der Kontrollbedingung konfluieren die beiden Aktivitätsschwerpunkte an der Mittellinie. Bei Ausschaltung callosaler Eingänge nehmen die Korrelationen mit der Idealverteilung ab und die Repräsentationen in den beiden Arealen schrumpfen.
Dies wird auch durch Messungen unterschwelliger lokaler Feldpotenziale gestützt, die ebenfalls auf eine Rolle callosaler Verbindungen bei großen und räumlich summierenden Reizen hinweisen [7].
Zukünftige elektrophysiologische Untersuchungen sollten darauf abzielen, die verschiedenen Projektionen, die in die Vermittlung des Kontextes involviert sind, aufgrund ihrer Dynamik zu entschlüsseln.
Originalveröffentlichungen
1.
J. Niessing, B. Ebisch, K. E. Schmidt, M. Niessing , W. Singer, R. A. W. Galuske:
Hemodynamic signals correlate tightly with synchronized gamma oscillations.
Science 309, 948-951 (2005).
2.
A. Thiele, G. Stoner:
Neuronal synchrony does not correlate with motion coherence in cortical area MT.
Nature 32, 366-370 (2003).
3.
K. E. Schmidt, M. Castelo-Branco, R. Goebel, B. R. Payne, S. G. Lomber, R. A. W. Galuske:
Pattern motion selectivity in population responses of area 18.
European Journal of Neuroscience 24, 2363-2374 (2006).
4.
K. E. Schmidt, R. Goebel, M. Castelo-Branco, S. G. Lomber, B. R. Payne, R. A. W. Galuske:
Global motion representation in primary visual cortex of the cat and the influence of feedback.
European Journal of Neuroscience 12 (Supplement 11), 39.15 (2000).
5.
Y. Egenolf, R. A. W. Galuske, S. G. Lomber, W. Barnes, B. R. Payne, W. Singer, K. E. Schmidt:
Impact of pMS feedback connections on the representation of contextual stimuli in cat area 18.
FENS Abstracts 2, A017.5 (2004).
6.
T. Wunderle, W. Barnes, N. Kriegeskorte, A. Schmidt, K. E. Schmidt:
Impact of visual callosal connections on the representation of center and surround stimuli in cat area 17 and 18.
Society for Neuroscience Abstracts 33, 920.16 (2007).
7.
V. A. Makarov, K. E. Schmidt, N. P. Castellanos, L. Lopez-Aguado, G. M. Innocenti:
Stimulus-dependent interaction between the visual areas 17 and 18 of the 2 hemispheres of the ferret (Mustela putorius).
Cerebral Cortex, Dec 7 [Epub ahead of print] (2007).
, ihre beiden Komponenten (G, H) und ein Modellstimulus für ihre Musterbewegung (C). Aktivitätskarten wurden entweder durch die Summe der beiden Komponentenaktivierungen (erste Spalte) oder die Mustermodelaktivität (zweite Spalte) dividiert. Ein Plaid mit niedrigem Schnittstellenkontrast (A, E), bevorzugt als zwei Ebenen wahrgenommen, ruft Aktivitätsverteilungen hervor, die durch die Summe der beiden Komponentenaktivierungen erklärt werden können (flache Modulation in A, starke komponentenähnliche Modulation in E, vergleiche E, G, H). Ein Plaid mit hohem Schnittstellenkontrast wird meist als sich in die Gesamtrichtung bewegendes Muster wahrgenommen. Tatsächlich aktiviert das Plaid in der primären Sehrinde Domänen, die denen des Mustermodells ähneln (starke Musteraktivierung in B, flache komponentenähnliche Modulation in F, vergleiche B, C), obgleich einzelne Neurone nur kleine Ausschnitte des Gesamtplaids bearbeiten. D: Oberflächenbild des abgeleiteten Gebiets. Rote Sterne in den Mustermodelldomänen (C) dienen der Orientierung. Die Antwortstärke ist als Grauwert kodiert.")

, bei pMS-Kühlung (blau) und nach Abschalten der Kühlung (grau). Je stärker die Aktivierung, desto heller die Farbe. Es fällt auf, dass im Gegensatz zum schwach kontrastierten Plaid (erste Spalte) das Plaid mit hohem Schnittstellenkontrast (zweite Spalte) die gleichen Domänen (gelb) aktiviert wie das Mustermodell (dritte Spalte). Bei pMS-Kühlung nimmt die Musteraktivität für dieses Plaid ab, wohingegen sie für das Modell erhalten bleibt. Dafür nimmt die komponentenähnliche Aktivität zu. Alle Effekte verschwinden bei Erwärmung von pMS wieder.")

 und Ganzfeld (dritte Spalte) vertikaler Orientierung während Kühlung des Areals pMS. Sterne kennzeichnen die Zentren der aktiven Domänen bei Ganzfeldstimulation in der Kontrollbedingung (oben rechts, rot: Überlappung mit der Ganzfeldkontrolle, gelb: Nichtüberlappung). A: Aktive Domänen in der Kontrollbedingung. Erste Spalte: Umfeldaktivierung vornehmlich anterior, zweite Spalte: Feldaktivierung vornehmlich posterior. Trotz nicht überlappender Sehreize werden einige Domänen von sowohl Umfeld- als auch Feldstimuli angesprochen (rote Sterne links der weißen Linie). B: Bei pMS-Deaktivierung nimmt die Antwortstärke für alle Reize ab. Dabei schrumpfen die vom Umfeld modulierten Bereiche sowie die Feld/Umfeld-Überlappungszone. C: Erholungsphase. D: Kortexoberfläche. E: Kühleffekte wirken sich stärker aus auf Umfeld- als auf Feldreizung: Der prozentuale Abfall der Übereinstimmung mit der Ganzfeldkontrolle ist bei Umfeldreizung signifikant (n=13). Die Antwortstärke ist als Grauwert kodiert.")