Dynamik und Plastizität der Informationsverarbeitung in der Großhirnrinde
Forschungsbericht (importiert) 2005 - Max-Planck-Institut für Hirnforschung
Autoren
Galuske, Ralf A. W.
Abteilungen
Neurophysiologie (Prof. Dr. Wolf Singer) MPI für Hirnforschung, Frankfurt/Main
Zusammenfassung
Die Signalverarbeitung in der Großhirnrinde wird maßgeblich von den zugrunde liegenden Verbindungsstrukturen bestimmt. Bisherige Konzepte dazu bauen im Wesentlichen auf der Rolle von feed-forward-Verbindungen auf, die primäre und höhere kortikale Areale verbinden. Dieser Ansatz vernachlässigt jedoch, dass auch in der Gegenrichtung massive Projektionen Signale von höheren Arealen zurück in die primären sensorischen Hirnareale bringen. Wir haben die funktionelle Rolle solcher feed-back-Verbindungen durch Kombination von optischen und elektrophysiologischen Ableitemethoden mit reversiblen Deaktivierungsmethoden untersucht und konnten zeigen, dass feed-back -Verbindungen eine wichtige Rolle bei der Verarbeitung von sensorischen Eingangssignalen spielen. Des Weiteren wurde untersucht, wie erfahrungsabhängige Plastizität von kortikalen Repräsentationen gesteuert wird. Diese Studien ergaben, dass die Induktion von plastischen Veränderungen in kortikalen Netzwerken entscheidend vom Kontext und globalen Aktivierungszustand, in dem die entsprechenden Reize präsentiert werden, abhängen.
Integration von feed-forward- und feed-back- Signalen im visuellen Kortex
In frühen sensorischen Arealen der Grosshirnrinde sind viele der Eigenschaften der zu verarbeitenden Information in so genannten „Karten“ in kolumnärer Weise repräsentiert. In den frühen kortikalen Verarbeitungsarealen des visuellen Systems bilden Eigenschaften wie z.B. die Orientierung von Konturen oder die Bewegungsrichtung von visuellen Reizen die Basis dieser komplexen Repräsentationen (Abb. 1).
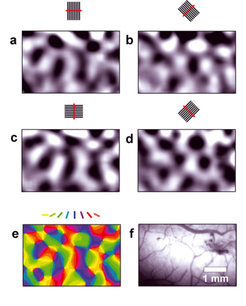
Orientierungskarten im primären visuellen Kortex. a) –d) Optisch abgeleitete Aktivitätsmuster im primären visuellen Kortex nach Stimulation mit orientierten Balkengittern. Die entsprechenden Stimuli sind jeweils oberhalb der Aktivitätskarten gezeigt. Dunkele Bereiche zeigen aktive Regionen, helle Bereiche zeigen inaktive Regionen. Ein Bild des abgeleiteten Bereiches ist in f) gezeigt. e) Farbkodierte Orientierungspräferenzkarte für diesen Bereich. Die Orientierungspräferenzen für jeden Pixel sind durch vektorielle Addition der in a)-d) gezeigten Einzelaktivitäten berechnet und dann gemäß des über der Abbildung angegebenen Farbschemas kodiert worden.
Orientierungskarten im primären visuellen Kortex. a) –d) Optisch abgeleitete Aktivitätsmuster im primären visuellen Kortex nach Stimulation mit orientierten Balkengittern. Die entsprechenden Stimuli sind jeweils oberhalb der Aktivitätskarten gezeigt. Dunkele Bereiche zeigen aktive Regionen, helle Bereiche zeigen inaktive Regionen. Ein Bild des abgeleiteten Bereiches ist in f) gezeigt. e) Farbkodierte Orientierungspräferenzkarte für diesen Bereich. Die Orientierungspräferenzen für jeden Pixel sind durch vektorielle Addition der in a)-d) gezeigten Einzelaktivitäten berechnet und dann gemäß des über der Abbildung angegebenen Farbschemas kodiert worden.
Derzeitige Konzepte zur Verarbeitung von sensorischen Informationen basieren auf der Annahme, dass verschiedene subkortikale Zentren und kortikale Areale die eingehenden Signale in hierarchischer Reihenfolge sequenziell analysieren und so die Grundlage für die kartenartigen Repräsentationen bilden. Eine solches, als „bottom-up“ bezeichnetes Verarbeitungskonzept vernachlässigt allerdings die Tatsache, dass neben den in der Hierarchie vorwärts (feed-forward) gerichteten Verbindungen auch ein noch dichteres Netzwerk von Rückprojektionen (feed-back-Verbindungen) besteht, durch das hierarchisch höher stehende Areale Signale zurücksenden können. Wir haben in den vergangenen Jahren begonnen, die physiologischen Eigenschaften dieser Rückprojektionen näher zu charakterisieren. Als Modell dient uns das visuelle System von Katzen. Ähnlich wie im visuellen System von Primaten lassen sich hier zwei getrennte Verarbeitungspfade grob unterscheiden, die Form und Bewegung getrennt in einer Kette von reziprok miteinander verbundenen Hirnarealen mit zunehmender Komplexität verarbeiten.
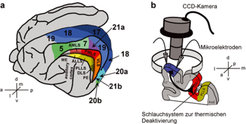
Schematische Darstellung des Versuchsaufbaus zur Untersuchung des Einflusses von feed-back- Verbindungen auf die neuronale Verarbeitung im primären visuellen Kortex. a) Seitenansicht des Großhirns der Katze mit Darstellung der verschiedenen visuellen Areale (modifiziert nach Tusa et al. 1981 [4]). b) Schematischer Versuchaufbau zur Untersuchung des Einflusses von feed-back-Verbindungen von Area PMLS nach Area 18, eines Teiles des primären visuellen Kortex bei der Katze. Die Mikroelektroden wurden zur elektrophysiologischen und die CCD-Kamera über Area 18 zur optischen Messung der neuronalen Aktivität benutzt. Die Orientierung der beiden Abbildungen ist durch die Richtungskreuze angezeigt (l: lateral, m: medial, d: dorsal, v: ventral, a: anterior, p: posterior).
Schematische Darstellung des Versuchsaufbaus zur Untersuchung des Einflusses von feed-back- Verbindungen auf die neuronale Verarbeitung im primären visuellen Kortex. a) Seitenansicht des Großhirns der Katze mit Darstellung der verschiedenen visuellen Areale (modifiziert nach Tusa et al. 1981 [4]). b) Schematischer Versuchaufbau zur Untersuchung des Einflusses von feed-back-Verbindungen von Area PMLS nach Area 18, eines Teiles des primären visuellen Kortex bei der Katze. Die Mikroelektroden wurden zur elektrophysiologischen und die CCD-Kamera über Area 18 zur optischen Messung der neuronalen Aktivität benutzt. Die Orientierung der beiden Abbildungen ist durch die Richtungskreuze angezeigt (l: lateral, m: medial, d: dorsal, v: ventral, a: anterior, p: posterior).
Um die Rolle von feed-back-Signalen zu untersuchen, haben wir höhere Areale im parietalen Pfad, der sich vornehmlich mit der Verarbeitung von Bewegungsinformationen befasst, durch Kühlung reversibel deaktiviert und gleichzeitig mithilfe optischer und elektrophysiologischer Ableitemethoden visuell evozierte Aktivität im primären visuellen Kortex untersucht (Abb. 2). Ein wesentliches Ergebnis dieser Experimente ist, dass die Deaktivierung dieser höheren Areale deutliche Aktivitätsverringerungen in frühen visuellen Arealen bewirkt. Zusätzlich zeigte sich, dass die Repräsentationen von Bewegungen nicht nur schwächer, sondern auch qualitativ verändert werden (Abb. 3). Insbesondere sehr bewegungsselektive Neurone veränderten in Abwesenheit von feed-back-Signalen ihr Feuerverhalten sehr stark bis hin zum völligen Erliegen der Antworttätigkeit.
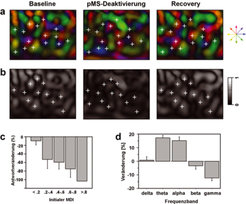
Konsequenzen der Deaktivierung von PMLS, eines auf die Verarbeitung von Bewegungsinformationen spezialisierten Kortexareales, auf Repräsentationen und neuronale Aktivität in Area 18. a) Farbkodierte Richtungskarten in Area 18 vor (links), während (mitte) und nach (rechts) der Deaktivierung von PMLS. Die weißen Kreuze markieren jeweils dieselben Orte in den drei Karten; die Zuordnung der verschiedenen Farben zu den unterschiedlichen Bewegungsrichtungen ist rechts außen angezeigt. Es fällt auf, dass während der PMLS-Deaktivierung die Zuordnung der verschiedenen richtungsselektiven Bereiche deutlich verändert ist. b) Karten der Vektorstärke, korrespondierend zu den Richtungskarten in a). Helle Bereiche weisen eine hohe Vektorstärke auf, dunkle Bereiche eine geringe. Während der Deaktivierung von PMLS (mittlere Karte) ist die Vektorstärke der visuell evozierten Antworten deutlich reduziert. c) Antwortveränderungen von Neuronen in Area 18 während der Deaktivierung von PMLS, gruppiert nach dem Ausmaß ihrer initialen Richtungsselektivität. Neuronen mit einer hohen Selektivität für bestimmte Bewegungsrichtungen weisen einen hohen MDI (mean direction index) auf, Neurone mit einer geringen Selektivität für eine bestimmte Bewegungsrichtung besitzen einen geringen MDI. Es fällt auf, dass insbesondere Neurone mit hohem MDI durch die Deaktivierung von feed-back- Verbindungen ihre Antwortstärke deutlich reduzieren. d) Veränderungen in der oszillatorischen Aktivität der neuronalen Antworten in Area 18 durch Deaktivierung von PMLS. Insbesondere schnelle Oszillationen im γ-Bereich sind vom Ausfall von PMLS betroffen.
Konsequenzen der Deaktivierung von PMLS, eines auf die Verarbeitung von Bewegungsinformationen spezialisierten Kortexareales, auf Repräsentationen und neuronale Aktivität in Area 18. a) Farbkodierte Richtungskarten in Area 18 vor (links), während (mitte) und nach (rechts) der Deaktivierung von PMLS. Die weißen Kreuze markieren jeweils dieselben Orte in den drei Karten; die Zuordnung der verschiedenen Farben zu den unterschiedlichen Bewegungsrichtungen ist rechts außen angezeigt. Es fällt auf, dass während der PMLS-Deaktivierung die Zuordnung der verschiedenen richtungsselektiven Bereiche deutlich verändert ist. b) Karten der Vektorstärke, korrespondierend zu den Richtungskarten in a). Helle Bereiche weisen eine hohe Vektorstärke auf, dunkle Bereiche eine geringe. Während der Deaktivierung von PMLS (mittlere Karte) ist die Vektorstärke der visuell evozierten Antworten deutlich reduziert. c) Antwortveränderungen von Neuronen in Area 18 während der Deaktivierung von PMLS, gruppiert nach dem Ausmaß ihrer initialen Richtungsselektivität. Neuronen mit einer hohen Selektivität für bestimmte Bewegungsrichtungen weisen einen hohen MDI (mean direction index) auf, Neurone mit einer geringen Selektivität für eine bestimmte Bewegungsrichtung besitzen einen geringen MDI. Es fällt auf, dass insbesondere Neurone mit hohem MDI durch die Deaktivierung von feed-back- Verbindungen ihre Antwortstärke deutlich reduzieren. d) Veränderungen in der oszillatorischen Aktivität der neuronalen Antworten in Area 18 durch Deaktivierung von PMLS. Insbesondere schnelle Oszillationen im γ-Bereich sind vom Ausfall von PMLS betroffen.
Die Ergebnisse dieser Experimente zeigen, dass feed-back-Signale eine essenzielle Rolle bei der Signalverarbeitung in frühen visuellen Arealen spielen: Zum einen wurde durch die Deaktivierung von höheren, Bewegungsinformation verarbeitenden Arealen im parietalen Kortex global die visuell evozierte Aktivität deutlich reduziert, zum anderen wurde die Repräsentation von Bewegungsrichtungen, die so genannte Richtungskarte, massiv verändert (Abb. 3). Feed-back-Signale sind somit in zweierlei Hinsicht sehr wichtig für die Verarbeitung von einlaufenden Informationen: Einerseits wird durch diese Signale das Aktivitätsniveau global erhöht und dies führt zur Verstärkung der Eingangssignale; andererseits kontrolliert die feed-back-Aktivität sehr spezifisch die Bewegungssensitivität der Neurone und formt damit die entsprechenden Repräsentationen, wodurch eine bessere Analyse der einlaufenden Signale ermöglicht wird. Dieser Effekt der feed-back-Aktivität kommt insbesondere bei der Analyse komplexer Stimuluskonfigurationen zum Tragen, deren Interpretation die Koordination der Aktivität von Neuronenverbänden erfordert, die über eine große Fläche verteilt liegen. Diese Koordination der Aktivität scheint sich in der zeitlichen Struktur von Aktivitätsmustern auszudrücken. Wir konnten beobachten, dass durch Wegfall der feed-back-Aktivität die präzise Synchronisation der neuronalen Antworten in primären visuellen Arealen reduziert wird und die damit einhergehende Feldpotenzial-Aktivität im hochfrequenten γ-Bereich (30-70 Hz) signifikant abnimmt (Abb. 3). Diese Daten legen nahe, dass die neuronalen Prozesse, die der Wahrnehmung zugrunde liegen, nicht nur auf der seriellen Verarbeitung einlaufender Informationen beruhen, sondern dass die lokalen Prozesse der globalen Kontrolle von hierarchisch höhergestellten Großhirnbereichen unterworfen sind und dadurch Teilinformationen nach globalen Ordnungskriterien zusammengefügt werden. Laufende Untersuchungen setzen an dieser Stelle an und versuchen herauszustellen, wie diese Interaktionen im Detail ablaufen und welche Faktoren die Aktivitätsmuster in höheren Arealen der Großhirnrinde beeinflussen.
Plastizität sensorischer Repräsentationen im visuellen Kortex
Nicht nur der auf einer sehr engen Zeitskala ablaufende Prozess der Interaktion von feed-back- und feed-forward-Interaktionen ist wichtig für die Verarbeitung von Sinnesinformationen, sondern auch der jeweilige Zustand der verarbeitenden Neuronennetze. Dieser Zustand kann in erheblichem Umfang von vorhergehenden Wahrnehmungen abhängen, da diese zu plastischen Veränderungen führen können, die als Lernprozesse in Erscheinung treten. In der Wahrnehmungspsychologie ist bekannt, dass die wiederholte Konfrontation mit demselben Stimulus eine Verbesserung (Sensibilisierung) oder eine Verschlechterung (Adaptation) der Wahrnehmung des jeweiligen Stimulus nach sich ziehen kann -so stellt sich die Frage nach den neuronalen Prozessen, die bei wiederholter Stimuluspräsentation zu Funktionsänderungen führen und welche Faktoren diese Veränderungen bestimmen. Wir formulierten die Hypothese, dass der allgemeine Aktivierungszustand für diese Prozesse eine maßgebliche Rolle spielt. Dieser wird entscheidend durch die Aktivität von Hirnstammzentren, insbesondere der Formatio Retikularis, gesteuert, die indirekt über den Neuromodulator Acetylcholin die globale Aktivität im Neokortex beeinflusst und auf diese Weise die oszillatorische Aktivität und die zeitliche Struktur von neuronalen Antworten stark verändern kann. In Phasen der Ruhe oder im Schlaf ist die Aktivität dieser Hirnstammstruktur gering und die oszillatorische Aktivität im Kortex liegt im Bereich von niedrigen Frequenzen (1-10Hz). In Phasen der Wachheit und des aktiven Verarbeitens von Sinnesinformationen ist diese modulierende Aktivität deutlich erhöht und es kommt zu schnellen Oszillationen im γ-Bereich (30-70 Hz). Gleichzeitig werden die Entladungsmuster der kortikalen Neurone mit einer Präzision im Millisekundenbereich synchronisiert. Um unsere Hypothese zu testen, haben wir die wiederholte Präsentation von visuellen Reizen mit elektrischer Reizung der Formatio Retikularis gepaart. Dabei zeigte sich, dass sich die Repräsentationen der wiederholt gezeigten Stimuli deutlich vergrößerten, während eine Abnahme der Signalstärke für diese Stimuli zu verzeichnen war, wenn die visuellen Reize ohne die Hirnstammaktivierung wiederholt dargeboten wurden (Abb. 4). Diese Effekte waren sehr eng an das Vorliegen von γ-Oszillationen gekoppelt, und auch auf dem Niveau von einzelnen Neuronen ließ sich nachweisen, dass sich die Antworteigenschaften der Zellen entsprechend veränderten: Fand die wiederholte Präsentation der Stimuli in Phasen erhöhter γ-Aktivität statt, so antworteten die Neuronen im Folgenden besser auf diese Stimuli; war die γ-Aktivität nicht erhöht, so antworteten die Zellen später schlechter auf die wiederholt gezeigten Reize.
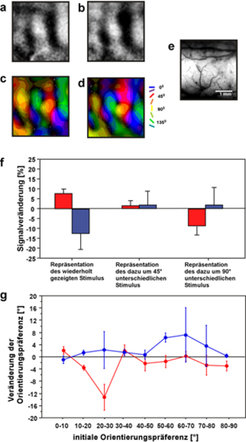
Veränderungen von Orientierungskarten nach wiederholter Stimulation mit orientierten Reizen: a) Orientierungskarte vor wiederholter Stimulation. b) Orientierungskarte nach wiederholter Stimulation; die Präsentation des visuellen Reizes wurde jeweils mit einem kurzen elektrischen Impuls in der Formatio Retikularis gepaart. c) und d) Farbkodierte Orientierungspräferenzkarten von derselben Ableitregion wie in a) und b) vor (c) und nach (d) repetitiver visueller Stimulation. Der wiederholt gezeigte Reiz hatte eine Orientierung von 45°, die in diesen Karten in rot kodiert ist. Man sieht die deutlichen Ausbreitungen der Repräsentation dieses Stimulus in die Randbereiche der entsprechenden Orientierungsdomänen. e) Oberflächenaufnahme von der in a)-d) abgeleiteten Region in Area 18. f) Mittlere optische Signalveränderungen durch wiederholte Stimulation mit demselben Stimulus mit und ohne gleichzeitige Hirnstammreizung in den verschiedenen Bereichen der Orientierungsrepräsentationen. Rote Balken zeigen die Werte für Fälle, in denen visuelle und elektrische Stimulation gepaart waren, blaue Balken zeigen Werte für Fälle, in denen nur visuell stimuliert wurde. Man sieht, dass in Fällen gepaarter Stimulation die Antwortstärke für den präsentierten Stimulus ansteigt, während bei alleiniger visueller Stimulation die Antworten schwächer werden. g) Mittlere Veränderungen der Orientierungspräferenz von elektrophysiologisch abgeleiteten Neuronen in Area 18. Die initiale Orientierungspräferenz der Neurone ist als Funktion ihrer Entfernung von der Orientierung des wiederholt gezeigten Stimulus aufgetragen. Die rote Kurve zeigt die Veränderungen der Neurone, die visuell in Phasen erhöhter γ-Aktivität stimuliert worden sind, während die blaue Kurve Veränderungen in Zellen zeigt, die in Phasen erniedrigter γ-Aktivität stimuliert wurden. Man sieht, dass Neurone, deren Orientierungspräferenz bis zu 30° unterschiedlich vom wiederholt gezeigten Reiz ist, ihre Orientierungspräferenz verändern, wenn der Reiz in Phasen erhöhter γ-Aktivität präsentiert wird und somit in Folge besser auf diesen Reiz antworten können.
Veränderungen von Orientierungskarten nach wiederholter Stimulation mit orientierten Reizen: a) Orientierungskarte vor wiederholter Stimulation. b) Orientierungskarte nach wiederholter Stimulation; die Präsentation des visuellen Reizes wurde jeweils mit einem kurzen elektrischen Impuls in der Formatio Retikularis gepaart. c) und d) Farbkodierte Orientierungspräferenzkarten von derselben Ableitregion wie in a) und b) vor (c) und nach (d) repetitiver visueller Stimulation. Der wiederholt gezeigte Reiz hatte eine Orientierung von 45°, die in diesen Karten in rot kodiert ist. Man sieht die deutlichen Ausbreitungen der Repräsentation dieses Stimulus in die Randbereiche der entsprechenden Orientierungsdomänen. e) Oberflächenaufnahme von der in a)-d) abgeleiteten Region in Area 18. f) Mittlere optische Signalveränderungen durch wiederholte Stimulation mit demselben Stimulus mit und ohne gleichzeitige Hirnstammreizung in den verschiedenen Bereichen der Orientierungsrepräsentationen. Rote Balken zeigen die Werte für Fälle, in denen visuelle und elektrische Stimulation gepaart waren, blaue Balken zeigen Werte für Fälle, in denen nur visuell stimuliert wurde. Man sieht, dass in Fällen gepaarter Stimulation die Antwortstärke für den präsentierten Stimulus ansteigt, während bei alleiniger visueller Stimulation die Antworten schwächer werden. g) Mittlere Veränderungen der Orientierungspräferenz von elektrophysiologisch abgeleiteten Neuronen in Area 18. Die initiale Orientierungspräferenz der Neurone ist als Funktion ihrer Entfernung von der Orientierung des wiederholt gezeigten Stimulus aufgetragen. Die rote Kurve zeigt die Veränderungen der Neurone, die visuell in Phasen erhöhter γ-Aktivität stimuliert worden sind, während die blaue Kurve Veränderungen in Zellen zeigt, die in Phasen erniedrigter γ-Aktivität stimuliert wurden. Man sieht, dass Neurone, deren Orientierungspräferenz bis zu 30° unterschiedlich vom wiederholt gezeigten Reiz ist, ihre Orientierungspräferenz verändern, wenn der Reiz in Phasen erhöhter γ-Aktivität präsentiert wird und somit in Folge besser auf diesen Reiz antworten können.
Diese Ergebnisse zeigen, dass der Kontext, in dem Reize verarbeitet werden, nicht nur eine wesentliche Rolle für die aktuelle Reizverarbeitung spielt, sondern dass er darüber hinaus die lang anhaltenden Veränderungen der verarbeitenden Strukuren nachhaltig bestimmt. Ferner zeigen diese Daten, dass die Induktion von plastischen Veränderungen in der Großhrinrinde ein zeitkritischer Prozess ist. In Phasen hoher γ-Aktivität sind die neuronalen Antworten im Millisekundenbereich synchronisiert und die Effizienz der synaptischen Übertragung zwischen den dann gleichzeitig aktiven Neuronen nimmt zu. In Phasen, in denen diese präzise Synchronisation nicht zustande kommt, werden die synaptischen Verbindungen abgeschwächt. Genau diese Veränderungen sind von dem kanadischen Psychologen Donald Hebb in den 1949 formulierten Lernregeln vorausgesagt worden. Unsere Experimente bestätigen diese Voraussage nunmehr auf systemischer Ebene.
Originalveröffentlichungen
1.
Galuske, R. A. W., K. E. Schmidt, R. Goebel, S. G. Lomber, and B. R. Payne:
The role of feedback in shaping neural representations in visual cortex.
Proceedings of the National Academy of Sciences USA 99, 17083-17088 (2002).
2.
Galuske, R. A. W.:
Reticular activation facilitates use-dependent plasticity in cat visual cortex.
European Journal of Neuroscience 12 (Suppl.11), 206.2 (2000).
3.
Hebb, D. O.:
The Organization of Behavior.
Wiley, New York (1949)
4.
Tusa, R. J., L. A. Palmer, and A. C. Rosenquist:
Multiple cortical visual areas: Visual field topography in the cat.
In: Woolsey, C. N. (ed.): Cortical sensory organisation, Vol. 2. Humana Press, Clifton, NJ, USA (1981).
 –d) Optisch abgeleitete Aktivitätsmuster im primären visuellen Kortex nach Stimulation mit orientierten Balkengittern. Die entsprechenden Stimuli sind jeweils oberhalb der Aktivitätskarten gezeigt. Dunkele Bereiche zeigen aktive Regionen, helle Bereiche zeigen inaktive Regionen. Ein Bild des abgeleiteten Bereiches ist in f) gezeigt. e) Farbkodierte Orientierungspräferenzkarte für diesen Bereich. Die Orientierungspräferenzen für jeden Pixel sind durch vektorielle Addition der in a)-d) gezeigten Einzelaktivitäten berechnet und dann gemäß des über der Abbildung angegebenen Farbschemas kodiert worden.")
 Seitenansicht des Großhirns der Katze mit Darstellung der verschiedenen visuellen Areale (modifiziert nach Tusa et al. 1981 [4]). b) Schematischer Versuchaufbau zur Untersuchung des Einflusses von feed-back-Verbindungen von Area PMLS nach Area 18, eines Teiles des primären visuellen Kortex bei der Katze. Die Mikroelektroden wurden zur elektrophysiologischen und die CCD-Kamera über Area 18 zur optischen Messung der neuronalen Aktivität benutzt. Die Orientierung der beiden Abbildungen ist durch die Richtungskreuze angezeigt (l: lateral, m: medial, d: dorsal, v: ventral, a: anterior, p: posterior).")
 Farbkodierte Richtungskarten in Area 18 vor (links), während (mitte) und nach (rechts) der Deaktivierung von PMLS. Die weißen Kreuze markieren jeweils dieselben Orte in den drei Karten; die Zuordnung der verschiedenen Farben zu den unterschiedlichen Bewegungsrichtungen ist rechts außen angezeigt. Es fällt auf, dass während der PMLS-Deaktivierung die Zuordnung der verschiedenen richtungsselektiven Bereiche deutlich verändert ist. b) Karten der Vektorstärke, korrespondierend zu den Richtungskarten in a). Helle Bereiche weisen eine hohe Vektorstärke auf, dunkle Bereiche eine geringe. Während der Deaktivierung von PMLS (mittlere Karte) ist die Vektorstärke der visuell evozierten Antworten deutlich reduziert. c) Antwortveränderungen von Neuronen in Area 18 während der Deaktivierung von PMLS, gruppiert nach dem Ausmaß ihrer initialen Richtungsselektivität. Neuronen mit einer hohen Selektivität für bestimmte Bewegungsrichtungen weisen einen hohen MDI (mean direction index) auf, Neurone mit einer geringen Selektivität für eine bestimmte Bewegungsrichtung besitzen einen geringen MDI. Es fällt auf, dass insbesondere Neurone mit hohem MDI durch die Deaktivierung von feed-back- Verbindungen ihre Antwortstärke deutlich reduzieren. d) Veränderungen in der oszillatorischen Aktivität der neuronalen Antworten in Area 18 durch Deaktivierung von PMLS. Insbesondere schnelle Oszillationen im γ-Bereich sind vom Ausfall von PMLS betroffen.")
 Orientierungskarte vor wiederholter Stimulation. b) Orientierungskarte nach wiederholter Stimulation; die Präsentation des visuellen Reizes wurde jeweils mit einem kurzen elektrischen Impuls in der Formatio Retikularis gepaart. c) und d) Farbkodierte Orientierungspräferenzkarten von derselben Ableitregion wie in a) und b) vor (c) und nach (d) repetitiver visueller Stimulation. Der wiederholt gezeigte Reiz hatte eine Orientierung von 45°, die in diesen Karten in rot kodiert ist. Man sieht die deutlichen Ausbreitungen der Repräsentation dieses Stimulus in die Randbereiche der entsprechenden Orientierungsdomänen. e) Oberflächenaufnahme von der in a)-d) abgeleiteten Region in Area 18. f) Mittlere optische Signalveränderungen durch wiederholte Stimulation mit demselben Stimulus mit und ohne gleichzeitige Hirnstammreizung in den verschiedenen Bereichen der Orientierungsrepräsentationen. Rote Balken zeigen die Werte für Fälle, in denen visuelle und elektrische Stimulation gepaart waren, blaue Balken zeigen Werte für Fälle, in denen nur visuell stimuliert wurde. Man sieht, dass in Fällen gepaarter Stimulation die Antwortstärke für den präsentierten Stimulus ansteigt, während bei alleiniger visueller Stimulation die Antworten schwächer werden. g) Mittlere Veränderungen der Orientierungspräferenz von elektrophysiologisch abgeleiteten Neuronen in Area 18. Die initiale Orientierungspräferenz der Neurone ist als Funktion ihrer Entfernung von der Orientierung des wiederholt gezeigten Stimulus aufgetragen. Die rote Kurve zeigt die Veränderungen der Neurone, die visuell in Phasen erhöhter γ-Aktivität stimuliert worden sind, während die blaue Kurve Veränderungen in Zellen zeigt, die in Phasen erniedrigter γ-Aktivität stimuliert wurden. Man sieht, dass Neurone, deren Orientierungspräferenz bis zu 30° unterschiedlich vom wiederholt gezeigten Reiz ist, ihre Orientierungspräferenz verändern, wenn der Reiz in Phasen erhöhter γ-Aktivität präsentiert wird und somit in Folge besser auf diesen Reiz antworten können.")